|
A |
B |
|
|
|
|
A |
B |
|
|
|
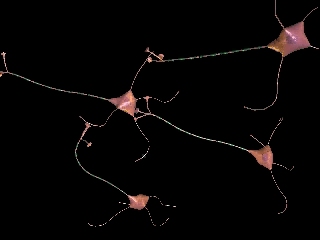
After release into the synaptic cleft, neurotransmitters interact with receptor proteins on the membrane of the postsynaptic cell, causing ionic channels on the membrane to either open or close. When these channels open, depolarization occurs, res ulting in the initiation of another action potential.
There are two types of postsynaptic receptors that recognize neurotransmitters. Ionotropic receptors, also referred to as ligand-gated ion channels, act quickly to depol arize the neuron and pass on the action potential (or hyperpolarize the neuron and inhibit additional action potentials). These receptors are made up of five individual protein subunits embedded in the cell membrane, and arranged to form a single pore th at spans this membrane. When a neurotransmitter associates with the extracellular recognition site, the membrane-spanning subunits of the receptor quickly open to form a pore through which the necessary ions can pass. Depolarization usually occurs a m illisecond or two after the action potential has been received and lasts only up to ten milliseconds.
?-aminobutyric acid(GABA) is one example of a neurotransmitter recognized by an ionotropic receptor. GABA is an inhibitory neurotransmitter used at roughly one-third of the synapses in the brain. The binding of GABA at the GABA recognition site c auses the membrane-spanning channel of the receptor protein to open and allow an influx of negatively charged chloride ions. This influx of negative ions serves to hyperpolarize the cell thus inhibiting the firing of an action potential. Though in the c ase of GABA, the ionotropic receptor is used to inhibit the firing of an action potential, there are other ionotropic receptors which recognize excitatory neurotransmitters and thus stimulate the firing of action potentials in post-synaptic cells. < /P>
Metabotropic receptors, or G-protein linked receptors, do not work as simply as ligand-gated ion channels do. Like ionotropic receptors, metabotropic receptors also have an extracellular neurotransmitter recognition site, yet these receptors do not form a membrane-spanning pore that can allow the direct passage of ions. Instead, when a neurotransmitter associates with the extracellular recognition site, an intermediate molecule within the postsynaptic cell, called a G-protein, is activated and, either directly or through a series of enzymatic reactions, opens or closes ion channels located at other places on the cell membrane. Because the action of metabotropic recepto rs is not as direct, their action is slower. Depolarization takes longer, typically lasting up to hundreds of milliseconds, and in some cases, going on for several minutes, hours, or even days.
Metabotropic receptors are used in the recognition of all neuropeptides and several small-molecule neurotransmitters. Dopamine (DA), for example, is a small-molecule neurotransmitter recognized by a G-protein coupled receptor. The binding of DA a t the recognition site of a post-synaptic DA receptor sets off a chain of reactions which ultimately cause ion pores along the post-synaptic membrane to open and an action potential to be stimulated. Found in four major tracts of the brain (the nigrostri al tract, the tuberoinfundibular tract, the mesolimbic tract, and the mesocortical tract), DA plays an important role in the control of both motor and emotional behavior.
A. Ionotropic (ligand-gated) receptors
|
|
1. GABAa Receptors: a representative family of ligand-gated receptors |
|
|
2. GABAa receptor function: inhibitory postsynaptic potentials |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
7. Positive neuromodulation of GABAa receptors: tranquilizers |
|
|
|
|
|
B. Metabotropic (G-protein coupled) receptors
|
|
1. Dopamine Receptors: a representative family of metabotropic receptors |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
© Williams College Neuroscience, 1998